|
E' noto come la variabilità animale (e vegetale) sia soggetta a selezione naturale e che quest' ultima rappresenti la principale forza deterministica del cambiamento evolutivo delle popolazioni naturali. La maggior parte dei biologi modemi ritiene, comunque, che vi siano anche altre cause dei cambiamenti evolutivi, altrettanto importanti, del tutto casuali e neutrali. Il Darwinismo classico già ammetteva l' esistenza di questi ultimi cambiamenti non riconoscendone, tuttavia, il rilevante peso e la notevole frequenza con cui si presentavano in natura. Oggi sappiamo che esistono due tipi variazioni evolutive: la "variazione casuale" ed il "cambiamento casuale"; la prima rappresenta il materiale grezzo su cui opera la selezione naturale per produrre cambiamenti nelle frequenze alleliche delle popolazioni; il secondo consiste in una modificazione direzionale delle stesse frequenze che si produce in piccole popolazioni in assenza di selezione, o meglio senza che la selezione giochi alcun ruolo su di esse. Gli effetti di queste ultime modificazioni sono del tutto casuali, non prevedibili: esse si originano, infatti, dal campionamento variabile del pool genico delle popolazioni naturali.
Il più noto esempio di cambiamento casuale e' rappresentato del "drift genetico", noto anche come deriva genetica, evoluzione neutrale o effetto Sewall Wright: esso consiste in un fenomeno casuale di oscillazione delle frequenze alleliche, strettamente correlato al numero di individui che costituiscono una popolazione. Il fenomeno, ben più frequente in natura di quanto un tempo si ammettesse, è interpretabile per mezzo del calcolo delle probabilità. Infatti è ben noto che se si lancia una moneta un numero di volte elevato, le probabilità di ottenere tante "croci" quante "teste" è alta ed aumenta con l'aumentare del numero di lanci effettuati. Al contrario, se si lancia la stessa moneta un numero basso di volte, ad esempio, 3, la probabilità di ottenere 3 teste o 3 croci consecutivarnente, cioè un risultato che si discosti da quello teorico, è maggiore. Secondo Mooto Kimura, autore della teoria della "neutralità selettiva" dell’evoluzione a livello della struttura del DNA e delle proteine, nella deriva genetica non c’è la sopravvivenza del più adatto bensì quella del più "fortunato". Molte popolazioni naturali subiscono grandi fluttuazioni nel numero di individui e sono sottoposte a periodi di massima espansione (aumento numerico di individui) e di contrazione (diminuzione numerica degli individui). In occasione di tali periodi, una popolazione naturale può diventare anche molto piccola e produrre un cosidetto effetto "a collo di bottiglia" (population bottle-neck). In tali circostanze, ossia nel caso di avere di fatto a che fare con una piccolissima popolazione, si può realizzare un effetto di "deriva genetica", cioè un cambiamento evolutivo indipendente dalla selezione naturale. 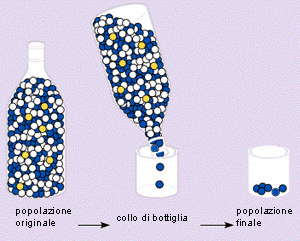 Consideriamo, ad esempio, una popolazione, di coleotteri che abbia subito un "collo di bottiglia" e che sia costituita da 10 individui dei quali 3 abbbiano un gene che produce antenne molto corte e che questi individui abbiano un 25% di vantaggio selettivo rispetto agli altri 7 individui. Supponiamo, altresi, che sfortunatamente un grosso animale, passando, nei pressi della popolazione, ne schiacci la metà, cioè 5 individui; in questo caso la probabilità che tra gli individui accidentalmente schiacciati ci siano proprio i 3 ad antenne corte, cioè quelli meglio adattati, risulta piuttosto elevata e consentirebbe come conseguenza la sopravvivenza degli individui meno adattati. Al contrario, qualora la popolazione fosse molto più numerosa, supponiamo 10.000 individui (3000 con antenne corte e 7000 con antenne normalmente sviluppate) la probabilità che lo stesso evento accidentale determini la morte di tutti i 3000 individui adattati sarebbe ovviamente molto minore. Possiamo, analogamente, prendere in considerazione una piccolissima popolazione di formiche, alcune delle quali con un particolare gene favorito dalla selezione, la quale venga a trovarsi accidentalmente su di un campo che improvvisamente bruci per una certa parte. Le probabilità che muoiano proprio le formiche con il gene favorevole è abbastanza alta; la stessa probabilità sarebbe, al contrario, bassa qualora la popolazione considerata fosse stata più numerosa. Allo stesso modo, l'uomo, potrebbe essere la causa indiretta di effetti di deriva genetica, operando frammentazione di areali e, quindi, di popolazioni, mediante incendi, sbarramenti di corsi d'acqua, costruzione di grandi autostrade, etc. Il "collo di bottiglia" rappresenta la manifestazione di cause e fattori di diversa origine. Le popolazioni naturali possono variare nelle loro dimensioni da una generazione all'altra a causa dell'avvicendarsi di stagioni più o meno favorevoli; in particolare il susseguirsi di un certo numero di annate sfavorevoli (siccità, etc.) può determinare effetti stocastici importanti (drift) nel pool genico delle relative popolazioni. Ad esempio in Groenlandia ed in Canada molte specie hanno potuto sopravvivere alle glaciazioni quaternarie in quanto sono state capaci di rifugiarsi nei "nunatack", cioè in quegli affioramenti di roccia circondati da ghiacciai perenni; in tale circostanza, comunque, le specie coinvolte hanno dovuto subire una drastica e stocastica riduzione, con conseguente imponente drift genetico di lunga durata, oltre 10.000 anni!
La perdita di variabilità genetica che si verifica a seguito di fenomeni di "collo di bottiglia" e di conseguente deriva genetica, dipende anche dalla velocità con cui le popolazioni che che lo hanno subito sono capaci di ripristinare le dimensioni originali; in ogni caso risulta poco significativa nel caso di un rapido recupero da parte della popolazione. Al contrario, popolazioni caratterizzate da ridotte dimensioni per un lungo periodo di tempo subiscono generalmente variazioni casuali nel loro pool genico e persino fissazione e perdita di alcuni alleli! In particolare popolazioni che risultino "cronicamente" piccole possono continuamente perdere geni e conseguentemente variabilità genetica: in definitiva la durata del "collo di bottiglia" gioca un ruolo determinante nella dinamica delle popolazioni naturali e nella loro variazione. Wright, ad esempio, ha stimato che una popolazione di ridotte dimensioni (20 individui) può raggiungere la condizione omozigote in circa 200 generazioni, una di 50 individui in 500, ed una di 1000 può conservare per lo stesso tempo una variabilità pari a circa il 95% di quella originale! Inoltre la durata del "collo di bottiglia" e la conseguente caduta di variabilità genetica può risultare sfavorevole soprattutto per quanto riguarda le capacità da parte di una popolazione di reagire ad eventi negativi, quale ad esempio un epidemia, etc. Un interessante esempio di deriva genetica è fornito da uno studio sui pesci che occupano sorgenti termali nella regione della Valle della Morte in California e in Nevada. Fatto notevole, una specie, Cyprinodon diabolis, è completamente confinata in una singola sorgente in Nevada, chiamata Devil’s Hole. Questa sorgente si formò circa 12.000 anni fa, dopo la fine dell’ultima glaciazione continentale, quando la regione fu coperta da un grande lago. Il Devil’s Hole è profondo oltre 60 m, ma i pesci sono in gran parte confinati in uno strato profondo soltanto circa 20 m, che conferisce a questi animali il più piccolo areale (area di distribuzione) che si conosca di tutte le specie di vertebrati. In realtà è possibile osservare simultaneamente ogni pesce. Il numero di individui nell’intera popolazione della specie varia nel tempo, ma spesso è soltanto di 50. Si ritiene che, a causa del piccolo ammontare della popolazione, la deriva genetica casuale sia stata importante nell’evoluzione di questa specie, che ha prodotto un pesce che è molto diverso dai pesci affini. L'importanza ed il particolare ruolo della deriva genetica e dell'effetto a "collo di bottiglia" nei processi evolutivi ha suscitato, comunque, numerosissime discussioni nel passato. Alcuni autori, ad esempio, hanno considerato già negli anni '30 questo effetto come una delle principali fonti della variabilità animale e vegetale e, quindi, dell'intera evoluzione biologica. Sempre negli stessi anni, e sino agli ultimi decenni, altri, tra cui Fischer, si fecero sostenitori di una corrente di pensiero del tutto opposta. Essi consideravano, infatti, tutta la variazione adattativa, nel senso che gli alleli delle diverse popolazioni dovrebbero avere ciascuno proprietà adattative diverse. Attualmente la scoperta che la gran parte delle popolazioni naturali sono polimorfe per enzimi che molto spesso differiscono poco nelle loro rispettive proprietà, ha riaperto e riacceso la discussione circa il vero significato e ruolo biologico di questo particolare aspetto dell'evoluzione biologica. Una forza stocastica molto simile a quella della deriva genetica è quella nota come "Effetto del Fondatore". Quando piccole o piccolissime popolazioni, o parte di una popolazione più grande, colonizzano un nuovo ambiente, e ciò può realizzarsi con diverse modalità (radiazione adattativa, frammentazione di areali, separazione di un' isola dal continente, trasporto passivo, etc.), i colonizzatori possono portare con se un campione incompleto del pool genico parentale. Nella nuova area colonizzata, pertanto, è possibile riscontrare individui portatori di geni rari, persino una certa frazione di eterozigosi, o addirittura geni disadattativi. Tale fenomeno, non a caso molto frequente nelle isole, laddove i processi di speciazione risultano molto intensi, viene, appunto, definito "effetto del fondatore" o "effetto del colonizzatore". 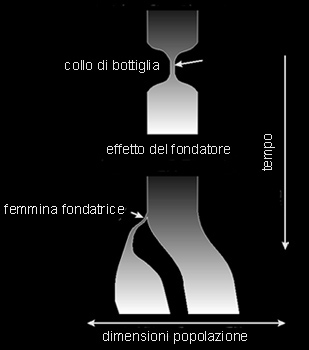 L'impostazione rigorosamente selezionista della scuola darwiniana accettava tale principio, come pure la deriva genetica, ma attribuiva a questi fenomeni un' importanza secondaria nei processi evolutivi, adducendo come giustificazione il fatto che in natura sarebbero poco frequenti le piccole popolazioni, presupposto indispensabile perchè i suddetti processi si possano realizzare. Oggi, al contrario, le nuove acquisizioni sul problema dell'evoluzione e gli studi approfonditi sulla struttura geografica delle specie, hanno inequivocabilmente dimostrato come la maggior parte delle specie siano suddivise in piccole o piccolissime popolazioni, spesso con scarsi contatti tra loro: tali popolazioni, o sottopopolazioni, a causa delle loro peculiari dimensioni e caratteristiche vengono attualmente considerate delle vere e proprie "unità evolutive". Ad esempio, se una specie di lepidotteri vive solamente sulle cime di montagne tanto distanziate tra loro che le farfalle non si possano spostare da una cima all'altra, anche se la popolazione può essere formata da un gran numero di individui, ciò evidentemente non ha alcuna importanza: infatti, ogni gruppo di farfalle che vive su una cima montuosa si comporterà indipendentemente dalle altre, costituendo di fatto una unità micropopolazionistica indipendente. Moltissimi sono gli esempi che potrebbero citarsi a proposito dell'effetto del fondatore in natura. E' oramai divenuto classico quello delle isole Bahams, molte delle quali attualmente ospitano ciascuna una forma differente di un gasteropode terrestre del genere Cerion, certamente non prodotte per mezzo di selezione naturale, bensì per un effetto del fondatore. Infatti, nel corso delle epoche glaciali, allorquando il livello del mare si abbassava a seguito dei ricorrenti cicli regressivi, alcune delle circa 700 isole erano in comunicazione tra loro e le chiocciole potevano spostarsi da un isola all'altra. Molte altre isole, al contrario, essendo circondate da mari più profondi, rimasero nello stesso periodo ancora separate tra loro; pertanto le chiocciole sono potute arrivare su queste ultime solo occasionalmente o accidentalmente, mediante trasporto passivo (su tronchi galleggianti, su uccelli, etc.), portando, con se una frazione molto particolare e generalmente piccola del pool genico delle popolazione originaria. In alcuni casi tali popolazioni, essendo questi gasteropodi ermafroditi, spesso autogami, sono state fondate a partire da un solo individuo che portava con se un pool genico evidentemente molto ridotto, poco rappresentativo di quello della popolazione da cui si sarebbe separato. Una conseguenza molto importante dell'effetto del fondatore è, inoltre, la possibile diminuzione o perdita totale di eterozigosi nella nuove popolazioni; e questo sarebbe imputabile a fenomeni di "inbreeding" (inincrocio) tra i suoi componenti; una tale situazione puo' comportare, molto spesso, come risultato l'esaltazione di geni e di fenotipi particolari come pure può, essere causa di difetti e malformazioni somatiche. Un esempio molto noto e significativo a questo riguardo è quello fornitoci dall "Old Order Amish" della Pensilvania, negli Stati Uniti. In questo gruppo, oggi consistente in circa 17.000 individui, si riscontra, con elevata frequenza una particolare malformazione, cioè polidattilia e nanismo, dovuta all'azione di un gene presente in molti dei suddetti individui. Dalla data della sua fondazione, avvenuta nel 1700, in questa comunità sono stati riscontrati ben 61 casi di questa malformazione, un numero all'incirca uguale a quello complessivo dei casi riscontrati in tutto il mondo nello stesso periodo di tempo. La causa di tale fenomeno è stata certamente un effetto del fondatore. Infatti, l'intero gruppo è stato fondato da solo tre coppie di irlandesi trasferitesi negli Stati Uniti, uno dei quali per caso deve essere stato il portatore del gene in oggetto. Tale gene si sarebbe successivamente affermato a seguito di inincrocio e di omozigosi tra i membri di questa comunità, peraltro chiusa, per motivi religiosi, a eventuali matrimoni con membri estranei alla comunità stessa. In tempi abbastanza recenti è stata avanzata l'ipotesi che anche la colonizzazione degli ambienti sotterranei acquatici (falde idriche, laghi, ambienti interstiziali etc.) si sarebbe realizzata mediante effetti combinati di deriva genetica ed effetto del fondatore. E' noto, infatti, come questo processo coinvolge generalmente piccole popolazioni o frazioni di popolazioni più ampie preadattate le quali, a causa di forti pressioni selettive esterne sono "costrette" a migrare negli ambienti sotterranei. In ogni caso si tratta di piccoli o piccolissimi gruppi portatori di una minima e casuale frazione della variabilità della popolazione di origine. Recenti studi hanno anche dimostrato che una tale diminuzione di variabilità è strettamente limitata solo alla prima fase del processo di colonizzazione; al contrario, allorquando la popolazione si sarà stabilizzata nel nuovo ambiente, la sua variabilità tenderà ad aurnentare progressivarnente, sino a superare in alcuni casi quella delle corrispondenti popolazioni epigee. Quest' ultimo aspetto, apparentemente illogico, potrebbe trovare la sua giustificazione nel fatto che gli arnbienti sotterranei, sia acquatici che terrestri, già ritenuti poco variabili e "monotoni" da un punto di vista ambientale, risultano molto spesso costitutiti da una miriade di microhabitat che possono, evidentemente, esercitare pressioni selettive molto diverse sui relativi popolamenti.
All Rights Reserved. Text and images on this website may not be redistributed or put
|