|
Il modo piů efficace per impedire il flusso genico con una popolazione parentale, piů ampia, č che una piccola frazione di essa si separi geograficamente, isolandosi totalmente dalla popolazione d'origine: questo particolare processo evolutivo viene definito, appunto, "speciazione allopatrica". La separazione puň realizzarsi con modalitŕ diverse: per esempio, puň essere il risultato dell'emigrazione attiva di una piccola popolazione dall'areale della popolazione parentale, il risultato di un cambiamento di ambiente (modificazione di un corso d'acqua, regressioni e trasgressioni marine, impatto antropico, etc.) o la presenza di barriere fisiche quali catene montuose, deserti, oceani, etc. A seguito della separazione una popolazione puň sviluppare meccansimi di isolamento riproduttivo che impediscono gli incroci con individui della popolazione d'origine e con altre popolazioni: tali meccansimi possono essere precoci (pre-zigotici) o tardivi (post-zigotici) e possono riguardare sia la morfologia che la fisiologia ed il comportamento. Un caso molto particolare di isolamento spaziale č quello relativo ai cosidetti "isolati periferici", da molti autori considerati le unitŕ evolutive piů importanti nei processi di speciazione. Lo stesso Darwin, avendo intuito l'importanza di tali popolazioni, attribuiva ad esse il nome di "specie incipienti". Gli isolati periferici, infatti, si prestano in modo molto particolare ai rapidi cambiamenti evolutivi che possono comportare conseguente speciazione: essi, infatti, sono protetti dal flusso genico proveniente dal nucleo centrale della popolazione e sono per lo piů costituiti da piccoli o piccolissimi gruppi, spazialmente limitati, nei quali puň evidentemente giocare un ruolo molto importante la casualitŕ (deriva genica, effetto del fondatore, etc.). Gli isolati periferici, infine, si localizzano in ambienti caratterizzati da intensa attivitŕ selettiva per quei caratteri che li differenziano dal resto della popolazione: una specie, infatti, presenta un limite periferico proprio perchč al di la di questo l'ambiente le sarebbe ovviamente sfavorevole. Non tutti gli isolati periferici, tuttavia, possono essere considerati "specie nascenti"; infatti, la maggior parte di essi non evolve necessariamente in nuove specie, bensě segue il normale destino dei piccoli gruppi che vivono ai margini di una areale, cioč si riduce nel tempo sino, in molti casi, ad estinguersi del tutto. In particolare, secondo Luria et al. (1981) "per una piccola popolazione il fatto di costituire un isolato periferico equivale semplicemente a possedere il biglietto vincente di una lotteria: senza di esso non si puň vincere, ma i biglietti vincenti sono molto pochi", il che allude evidentemente alla scarse probabilitŕ di successo di una speciazione in queste condizioni. A tutt'oggi sono stati studiati centinaia di casi di speciazione allopatrica nelle isole, soprattutto in quelle dell'oceano Pacifico (Samoa, Tahiti, Marchesi, Hawai, Galapagos, etc.). Esempi divenuti ormai classici sono quelli che si riferiscono alle diverse specie di fringuelli e di tartarughe che vivono sulle isole Galapagos.
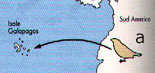
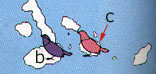
I fringuelli di Darwin rappresentano un esempio classico di speciazione allopatrica: le isole Galapagos si formarono nell'oceano Pacifico tra 3 e 5 milioni di anni fa. Situate a piů di 1000 Km Ovest dell'Ecuador, queste isole non sono mai state collegate alla terra ferma ed erano disabitate. Nonostante ciň, una piccola popolazione di fringuelli, che proveniva dal continente, ha colonizzato l'arcipelago e si č evoluta in specie diverse. Si puň ipotizzare che questa popolazione sia giunta nell'arcipelago durante una tempesta e si sia stabilizzata in una delle isole. Alcuni individui migrarono nelle isole piů vicine; in questo modo venne a crearsi tra loro una barriera geografica. Le diverse condizioni ecologiche nelle isole dell'arcipelago possono avere provocato variazioni significative nelle abitudini alimentari delle piccole popolazioni di fringuelli, riproduttivamente isolate. In qualche punto, pool genici di queste popolazioni isolate differivano talmente l'uno dall'altro da impedire l'interfecondazione. Le forme divergenti possono allora essere considerate specie diverse. Nel primo caso le 14 specie di fringuelli che si trovano attualmente su queste isole si sarebbero originate da una popolazione di fringuelli terricoli provenienti dal vicino continente i quali, separati geograficamente sulle diverse isole, avrebbero dato l'avvio ad un intenso processo di speciazione allopatrica; nel secondo caso le tartarughe rappresentano un esempio di analoga speciazione ancora in atto che, molto probabilmente, a causa della loro dissennata caccia da parte dell'uomo, non potrŕ forse mai concludersi! Un'altra interessante area di intensi processi di speciazione allopatrica čstato l'istmo di Panama, ripetutamente sommerso ed emerso a seguito dei numerosi cicli trasgressivi e regressivi del mare: a seguito di ciascuna emersione, gli oceani Pacifico ed Atlantico divennero "isole" per le popolazioni marine che, pertanto, andarono incontro a processi di isolamento e, quindi, di speciazione; al contrario, allorquando gli stessi oceani comunicavano tra loro a causa dell' abbassamento dell'istmo, furono i continenti che divennero a loro volta "isole" e centri di intensa speciazione. Numerosissimi sono ancora gli esempi che potrebbero essere citati: tra i piů significativi ricordiamo il Gran Caynon ed il corso del Colorado, insieme con la Baia di San Francisco, dove sono stati studiati interessanti casi di speciazione allopatrica in insetti, anfibi e mammiferi.
La speciazione simpatrica consiste nella formazione di nuove specie mediante isolamento riproduttivo all'interno dell'areale delle popolazioni parentali senza che intervenga isolamento geografico. Tale processo puň realizzarsi con tre differenti modalitŕ che vengono definite: speciazione simpatrica istantanea (o "quantum speciation"), speciazione simpatrica graduale e speciazione simpatrica catastrofica. Nel primo caso si forma istantaneamente, o comunque in tempi molto brevi, una barriera intrinseca al flusso genico tra alcuni individui ed il resto della popolazione; la causa di tale fenomeno potrebbe essere la comparsa improvvisa di una o piů mutazioni che limitino immediatamente lo scambio genico tra alcuni individui ed il resto della popolazione per cui i primi vengono a costituire un gruppo isolato riproduttivamente e geneticamente e, quindi, una nuova specie. Teoricamente questo tipo di speciazione, peraltro piuttosto rara negli organismi animali, si verifica allorquando gli individui portatori delle suddette mutazioni si accoppiano preferenzialmente tra di loro e se a favore di queste mutazioni agiscono pressioni selettive tali da controllare il flusso genico prodotto dagli incroci tra i mutanti ed il resto della popolazione. E' evidente che questo tipo di speciazione avrŕ tanto piů successo quanto piů piccola sarŕ la popolazione in cui si verifica: in questo caso, infatti, i nuovi cambiamenti non vengono "diluiti" in una popolazione ampia e costituita da individui "normali", bensě hanno buone probabilitŕ di diffondersi rapidamente mancando, o essendo molto ridotto l'effetto inerziale degli alleli della popolazione parentale. La maggior parte dei cambiamenti genetici, tuttavia, non conferisce un immediato isolamento riproduttivo: č questo il motivo per cui gran parte della speciazione simpatrica č graduale, si realizza cioč con l'instaurarsi dapprima di meccanismi isolanti deboli (post-zigotici) e, quindi, di meccanismi progressivamente piů efficienti (pre-zigotici). Tra questi ultimi, molto importanti sono quelli stagionali, ecologici, fisiologici ed etologici. Nel primo caso due popolazioni possono essere simpatriche, ma le loro stagioni riproduttive sono sfasate nel tempo sicchč i maschi di una non possono mai incrociarsi con le femmine dell'altra e viceversa; questo tipo di meccanismo isolante, molto comune nelle piante, risulta meno frequente tra gli animali (insetti, molluschi gasteropodi). In altri casi due popolazioni potenzialmente interfeconde tra loro non si riproducono in quanto utilizzano nicchie ecologiche diverse, sono ciočisolate ecologicamente. Particolarmente importanti ed efficaci sono anche i meccansimi isolanti fisiologici, soprattutto quelli che riguardano la biochimica dei gameti. Ad esempio nei comuni ricci di mare ed in altri animali a fecondazione esterna differenze biochimiche impediscono agli spermatozoi di una specie, o di una popolazione, di fecondare uova che non siano conspecifiche. Altri meccanismi particolarmente efficaci possono, infine, riguardare il corteggiamento: in molti uccelli, pesci ed insetti, infatti, sia i maschi che le femmine elaborano specifici segnali di riconoscimento sessuale (piumaggio, canto, movimenti, colorazioni, etc.) che impediscono l'ibridazione, favorendo altresěgli incroci intraspecifici ed intrapopolazionistici. La speciazione catastrofica, infine, si realizza in piccole popolazioni, ridotte drasticamente nelle dimensioni da catastrofi ambientali o da impatto antropico, con conseguente selezione di genotipi rari. Un altro particolare tipo di speciazione simpatrica čquello che si realizza molto frequentemente in specie parassite che, a seguito di mutazioni, possono cambiare la loro specificitŕ parassitaria; esse, cioč, perdono il contatto con la specie originaria, si riproducono preferenzialmente tra di loro e, come conseguenza dell'isolamento riproduttivo, possono evolvere in nuove specie. Quanto sinora esposto sulla speciazione simpatrica non č, comunque, condiviso da tutti gli autori: esistono, infatti, almeno tre evidenze in base alle quali la speciazione dovrebbe essere quasi sempre allopatrica. Da indagini di tipo storico risulta, infatti, che in tempi passati la speciazione č stata quasi sempre allopatrica. Da un punto di vista teorico risulta quasi impossibile che due specie affini possano coesistere in maniera ottimale nello stesso areale, utilizzando nicchie ecologiche identiche (principio di esclusione di Volterra-Gause). Per il mantenimento di due specie affini in condizioni simpatriche č necessario che queste non presentino sovrapposizioni di habitat e di nicchie ecologiche, soprattutto per quanto riguarda le preferenze alimentari e riproduttive: infatti, quando due o piů specie entrano in simpatria in generale si dividono habitat e nicchie e, pertanto, non vengono a competere tra di loro. La zoogeografia, infine, dimostra che specie molto affini sono generalmente localizzate in areali separati geograficamente e che negli stessi areali si riscontrano generalmente forme diverse, anche a livello generico.
Abbiamo visto come, sia nel caso della speciazione simpatrica che allopatrica, il blocco del flusso genico čconsiderato il requisito necessario e indispensabile affinchč si possa formare una nuova specie. Recentemente č stato, comunque, dimostrato che si puň realizzare un processo microevolutivo anche quando due popolazioni, pur separate geograficamente, continuano a mantenere un certo contatto spaziale e riproduttivo tra loro mediante una zona di confine: tali popolazioni vengono definite "parapatriche" ed il fenomeno cui possono dar luogo "speciazione parapatrica" (o stasipatrica). Casi comuni di speciazione parapatrica sono quelli che si riferiscono a popolazioni che divergono sino a livello di specie entro "clini" continui, senza l'intervento dell'isolamento geografico. Condizioni necessarie per cui il fenomeno abbia successo sono il mantenimento di un debole flusso genico tra le due popolazioni ed una diversa pressione selettiva ai due estremi del cline. Quest'ultimo fenomeno risulta molto frequente in animali con abitudini sedentarie nei quali lo sviluppo di meccanismi di isolamento riproduttivo accampagnano l'ingresso e l'adozione di nuovi habitat da parte di individui geneticamente divergenti. Casi di speciazione parapatrica sono stati messi in evidenza in numerosi gruppi ed in particolare nei lepidotteri, negli ortotteri ed in alcuni roditori con abitudini fossorie.
La speciazione filetica (o anagenesi) consiste nel fenomeno per cui una specie si trasforma in un'altra, che sostituisce la prima, seguendo un percorso lineare. Una singola specie puň trasformarsi gradualmente a tal punto da perdere i suoi principali connotati e trasformarsi in una nuova specie (speciazione successionale); in altri casi, una specie puň separarsi, sempre gradualmente, in due o piů linee divergenti, riproduttivamente isolate (speciazione divergente o cladogenesi). In ogni caso la formazione delle nuove entitŕ specifiche escludono la persistenza della specie d'origine: in questo caso, a differenza dei casi precedenti, il principale fattore d'isolamento č il tempo piuttosto che barriere biologiche o spaziali. Alcuni autori, per spiegare l'intensa e rapida speciazione in alcune specie di "Drosophila" delle Hawai, hanno proposto un altro modello di speciazione, intermedio tra quello simpatrico e quello allopatrico. Secondo questo modello, definito "Flush and Crash Evolution", il processo di speciazione prenderebbe l'avvio con l'arrivo in una nuova localitŕ (un isola) di pochissime femmine o di una sola femmina fecondata; la discendenza di tali femmine aumenterebbe rapidamente in una prima fase della colonizzazione (flush) a causa soprattutto della mancanza o scarsit ŕ di competitori e di una debole selezione naturale per cui, come conseguenza, il relativo pool genico della nuova popolazione puň cambiare e riorganizzarsi liberamente. Successivamente la popolazione subirebbe una forte contrazione (crash) a causa delle diminuite risorse trofiche dell'ambiente che funzionano ora quale intensa pressione selettiva sulla popolazione. Un tale modello di speciazione č molto simile a quello geografico in quanto prende l'avvio da un isolamento spaziale; d'altro canto č anche assimilabile a quello simpatrico istantaneo in quanto richiede tempi molto brevi per potersi realizzare. Molti autori, tuttavia, fanno rientrare questo peculiare modello di speciazione nell'effetto del fondatore. La "Flush and Crash Evolution" si realizza abbastanza frequentemente nelle diverse fasi di colonizzazione di isole, di altri habitat precedentemente non occupati da potenziali competitori e, sotto certi aspetti, anche nei processi di colonizzazione degli habitat acquatici ipogei da parte di specie dulciacquicole di superficie.
In questo tipo di speciazione una specie puň formarsi molto rapidamente, in alcuni casi anche in una sola generazione, sia attraverso effetti di deriva genica che di effetto del fondatore. Si conoscono due differenti tipi per questo tipo di speciazione: 1) un processo rapido di speciazione puň intervenire per ibridazione, quando due specie vengono a contatto e si riproducono; in tal caso viene prodotta una nuova specie in una sola generazione, Questo processo č piuttosto raro negli animali; 2) molte piante che si riproducono asessualmente, a seguito di poliploidia, possono generare nuove specie. Questo processo č sconosciuto tra gli organismi animali. Popolazioni simpatriche di specie strettamente affini possono produrre ibridi fertili e la popolazione che cosě si forma, mantenendosi nel tempo, acquisisce caratteristiche proprie che la separano dalle specie parentali (che nel contempo incrementano la loro divergenza) Tale modello di speciazione, apparentemente raro fra gli animali (č noto negli insetti fra i fasmidi e tra i vertebrati in Rana esculenta, prodotto dell' ibrido fra R. lessonae e R. ridibunda) č frequente nelle piante, dove si realizza per mezzo di allopoliploidia (da: Zunino & Zullini: Biogeografia. 2004).
LURIA S. E., S. J. GOULD & S. SINGER. 1989. Una visione della
vita. Introduzione alla biologia. HUXLEY J. 1974. Evolution. The modern synthesis. G. Allen & Unwin
Ltd., pp. 705.
MAYR E. 1970. L'evoluzione delle specie animali. G. Einaudi ed.,voll.I-II,
pp. 833 MITCHEL L. G., J. A. MUTCHMOR & W. D. DOLPHIN. 1992. Zoologia. Zanichelli Ed., pp. 1014
All Rights Reserved. Text and images on this website may not be redistributed or put
   |