|
L'intestino degli unirami č lungo ed in genere rettilineo, č suddiviso in intestino anteriore, medio, posteriore. Le ghiandole salivari sono associate ad una o pił appendici boccali. Nelle falene (bruchi), nelle api e vespe larvali, queste ghiandole secernono seta utilizzata per formare le cellule pupali. Gli unirami che si nutrono di cibi solidi producono una membrana peritrofica, questa serve a proteggere il delicato intestino medio dalle abrasioni e nell'intestino posteriore serve per incapsulare le feci in forma sferica. Negli insetti si sono evolute una serie di strutture specializzate per la digestione. L'intestino anteriore č suddiviso in faringe, esofago, ingluvie e proventricolo. L'ingluvie č un centro di immagazzinamento le cui pareti sono molto estensibili. Il proventricolo controlla il passaggio del cibo nell'intestino medio, presenta una valvola che separa i cibi semiliquidi oppure presenta un organo trituratore detto ventriglio che mastica i cibi solidi. L'intestino medio presenta dei ciechi gastrici posti nella zona di congiunzione con l'intestino anteriore, questi servono ad aumentare la superficie utile per l'assorbimento.
E' costituito da un cuore tubolare dorsale che pompa sangue verso il capo, questo organo si restringe anteriormente formando un'aorta vascolare da cui il sangue scorre posteriormente attraversando delle camere emoceliche per poi tornare nel seno pericardico e quindi al cuore passando dagli osti laterali. Spesso vi sono organi pulsanti alla base degli arti e delle ali. Tali organi aiutano la circolazione e la stabilitą della pressione sanguigna. L'attivitą pompante č debole e la pressione del sistema č bassa. Gli emociti nel sangue non hanno la funzione di trasporto dell'ossigeno, ma di cicatrizzazione e coagulazione.
Esistono strutture speciali per il trasporto dei gas respiratori, queste costituiscono il sistema tracheale, tale sistema permette gli scambi gassosi con l'ambiente senza correre il rischio della disidratazione. Le trachee sono delle estese invaginazioni tubulari della parete corporea, aperte attraverso la cuticola per mezzo di pori detti spiracoli. La parete di ogni trachea č sclerotizzata rinforzata da anelli detti tenidi. Queste trachee si dipartono dallo spiracolo e anastomizzano con altre a formare una rete che si estende in tutto il corpo. Gli spiracoli sono rientrati in un atrio le cui pareti sono rivestite da setole o tricomi che impediscono a polvere e parassiti di entrare nelle trachee. Gli spiracoli dei miriapodi sono circondati da un labbro sclerotizzato o peritrema con la stessa funzione dei tricomi. Il sistema tracheale č ventilato da gradienti di pressione indotti dall'animale stesso, lo spostamento d'aria č provocato dal movimento del corpo e dell'intestino. Alcuni insetti hanno regioni dette tasche tracheali che fungono da sacche per l'immagazzinamento dell'aria, l'attivitą muscolare di alcune regioni corporee contrae le sacche creando onde ventilatrici ritmiche.
Tali sacche si estendono accanto ad ogni organo o anche ad ogni fibra muscolare rifornendoli di ossigeno. Gli insetti acquatici hanno estroflessioni della parete corporea ricoperte da cuticola sclerotizzata contenenti sangue, trachee o sacche d'aria. Queste specie di branchie hanno canali che le congiungono al sistema tracheale principale.
La soluzione del problema dell'escrezione e della necessitą di limitare la perdita d'acqua č stata trovata nei tubuli malpighiani, questi sono escrescenze non ramificate dell'intestino nel tratto di congiunzione tra intestino medio e posteriore. Le estremitą distali cieche si estendono nell'emocele e sono poste tra i vari organi.
Il numero di tali strutture č estremamente variabile. Gli unirami sono rivestiti da una cuticola sclerotizzata con proprietą impermeabili che č un rimedio contro la disidratazione, soprattutto nei miriapodi, questi unirami sono costretti a vivere in ambienti umidi. Negli insetti č presente uno strato ceroso all'interno dell'epicuticola, tale strato aumenta la resistenza alla disidratazione e rende gli insetti maggiormente liberi di movimento e capaci di vivere anche in ambienti secchi. La filtrazione escretoria č spinta dalla pressione osmotica (poiché quella sanguigna č insufficiente).
Alcuni ioni vengono trasportati dal sangue attraverso l'epitelio del tubulo malpighiano nel suo lume mantenendo il gradiente osmotico che permette all'acqua di muoversi dalla cavitą corporea ai tubuli ed all'intestino. Vi sono delle regioni pił spesse contenenti speciali cellule dette ghiandole rettali che migliorano il processo di riassorbimento di acqua, sali, aminoacidi ed altre sostanze.
L'urato di potassio presente nei tubuli malpighiani viene precipitato in acido urico i cui cristalli non possono essere riassorbiti e pertanto vengono escreti con le feci. Negli insetti sono presenti anche particolari cellule dette nefrociti che si spostano nell'emocele digerendo le scorie.
Negli unirami il sistema nervoso č simile allo schema generale degli artropodi. I miriapodi presentano una scarsa fusione dei gangli, mentre č molto elevata e complessa negli insetti.
In molti insetti sono presenti anche delle fibre giganti. Come negli altri artropodi, i gangli cerebrali (cervello) comprendono tre zone distinte: protocerebro associato agli occhi, deutocerebro associato alle antenne, tritocerebro. Il ganglio subesofageo č composto dai gangli fusi del terzo, quarto e quinto segmento del capo e controlla le parti boccali, le ghiandole salivari e la muscolatura locale. Il ganglio ipocerebrale proprio degli insetti č posto tra il ganglio cerebrale e l'intestino anteriore; esistono due paia di corpi ghiandolari detti corpora cardiaca e allata associati al ganglio, questi funzionano in sintonia con le ghiandole protoraciche.
Il ganglio ipocerebrale ed i corpora costituiscono un complesso centro endocrino che regala prevalentemente la crescita e la metamorfosi. Gli unirami presentano degli ocelli semplici. Nei miriapodi sono gli unici occhi che si sviluppano, i centopiedi hanno molti ocelli che in alcune specie formano uno pseudo-occhio composto simile a quello di insetti e crostacei, ma č funzionale soltanto nel rilevamento della luce. I sinfili, i pauropodi e molti unirami parassiti non presentano strutture oculari. Negli insetti adulti gli ocelli formano una triade o una coppia sulla superficie antero-dorsale del capo.
La maggior parte degli insetti hanno due occhi composti che permettono un ampio campo visivo. Come gią detto nell'introduzione generale agli artropodi, l'occhio composto č formato da pił ommatidi, il cui numero determina la precisione visiva. 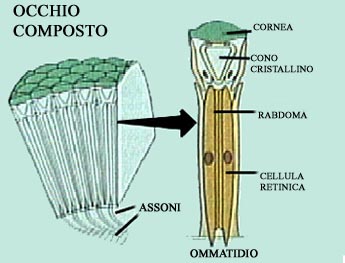
Ogni ommatidio č costituito da due elementi funzionali, la parte esterna comprende la cornea (lente) ed il cono cristallino, la parte interna comprende il rabdoma e le cellule retiniche. Quando gli occhi hanno un cono cristallino sono detti occhi euconi. Nella maggior parte degli insetti sotto la cornea sono presenti quattro cellule dette cellule Semper. Quando queste cellule non formano un cono cristallino e diventano trasparenti gli ommatidi sono detti aconi, tali ommatidi si ritrovano in alcuni emitteri, coleotteri e ditteri. In gran parte dei ditteri e degli odonati le cellule Semper producono coni pieni di liquido, gli ommatidi che hanno tali coni sono detti ommatidi pseudoconi. In alcuni coleotteri la lente si prolunga verso l'interno e le cellule Semper formano una struttura refrattaria tra cuticola e cellule retiniche, questi ommatidi sono detti esoconi. A differenza degli insetti, i miriapodi hanno l'organo di Tomosvary, questo č posto alla base delle antenne ed č formato da un disco con un poro centrale in cui convergono le estremitą dei neuroni sensoriali, tale organo costituisce un sensore chimico e un rilevatore di pressione, di umiditą e dei suoni. Come negli altri artropodi gli unirami hanno molti peli e setole sensoriali dette sensilli, sulla superficie corporea particolarmente concentrati sulle antenne, sulle appendici boccali e sulle zampe; i sensilli hanno funzione tattile o di sensori chimici. Spesso sulle antenne sono presenti setole sensoriali claviformi dette organi picchetto. Negli insetti sono presenti dei propriocettori interni detti organi cordotonali. In molti ordini di insetti vi sono molte strutture sensoriali diverse come i fonorecettori e gli organi timpanali.
Gli organi timpanali sono sull'addome, sul torace e sulle zampe anteriori. La comunicazione del suono negli insetti come la comunicazione luminosa č specie-specifico di comunicazione tra simili. Molti gruppi di insetti (es. ditteri, ortotteri ed emotteri) hanno strutture specifiche per la produzione di suoni utilizzate particolarmente per il corteggiamento. Le cicale hanno delle piastre od opercoli nella regione ventrale del metatorace, queste coprono un complesso sistema di membrane vibranti e camere risonanti, il suono esce da uno spiracolo metatoracico. Molti coleotteri hanno un meccanismo di comunicazione che sfrutta le increspature sulla superficie dell'acqua in modo simile al meccanismo dei ragni di rilevazione delle vibrazioni sulla ragnatela. Esistono anche organi che producono luce, questi sono costituiti da speciali cellule dette fotociti.
Negli unirami il comportamento riproduttivo č molto vario per i diversi taxon sia per quanto riguarda l'inseminazione (diretta e indiretta), la nascita (ovipara, ovovipara) e in alcuni insetti per la partenogenesi. Anche per lo sviluppo si riscontrano diversitą importanti: sviluppo diretto per i miriapodi e insetti primitivi (i piccoli sono "adulti in miniatura") sviluppo misto per insetti avanzati (larva, pupa, adulto). Chilopoda: i maschi e le femmine dei centopiedi posseggono genitali interni (ovaie e testicoli) che si aprono all'esterno al gonoporo, che si trova nel segmento genitale. L'inseminazione nei chilopodi č di tipo indiretto: il maschio deposita le spermatofore in una tela di fili setosi tessuti in una rete nuziale. Successivamente la femmina tramite i gonopodi (piccole appendici esterne del gonoporo) raccoglie la spermatofora e la inserisce nel proprio gonoporo, quindi deposita le uova sottoterra e nella vegetazione in decomposizione. Alcune specie nascono gią con tutti i segmenti , altri ne aggiungono con mute di passaggio dallo stato giovanile a quello allungato. Diplopoda: I millepiedi sia maschi che femmine posseggono un unico paio di gonadi allungate; i gonopori si aprono nel terzo segmento del tronco detto genitale. Nelle femmine ogni ovidotto si apre nell'atrio genitale (vulva), mentre nei maschi i dotti genitali terminano in uno o una coppia di peni. Nell'accoppiamento i peni e i gonopodi entrano in contatto, un enzima scioglie l'involucro delle spermatofore e gli spermatozoi si spostano nei ricettacoli seminali dei millepiedi femmina. Le uova si fecondano mentre vengono depositate sotto terra, o in un nido fatto con feci; vengono depositate anche 300 uova per volta. I piccoli nascono con solo sette segmenti e tre paia di zampe; ulteriori segmenti e zampe si aggiungono con mute successive. Pauropoda: il maschio deposita una spermatofora ad uno pił fili di seta tesa tra due pietre o foglie; la femmina le raccoglie e permette la fecondazione al suo interno. Da uova depositate sul legno decomposto nascono piccoli con pochi segmenti e con solo tre paia di zampe. Symphila: le spermatofore sono poste all'apice di steli; la femmina quando incontra questi pacchetti spermatici li morde ed immagazzina gli spermatozoi con cui feconda all'esterno ogni uovo. I piccoli nascono con circa la metą di segmenti dell'adulto. Insetti: i sessi sono separati come nei miriapodi, ma hanno un meccanismo di inseminazione di tipo diretto, la fecondazione č in genere interna. Le femmine hanno un paio di ovari composti da un grappolo di ovarioli tubulari i cui ovidotti formano un dotto comune che si immette nella camera genitale, anche i ricettacoli seminali o spermateche e le ghiandole annesse si aprono nella cavitą genitale che a sua volta si apre in una borsa copulatrice (gonoporo) sullo sternite dell'ottavo segmento addominale. I maschi hanno due testicoli con tubi spermatici che si appaiano per formare le vescicole seminali da cui si diparte il dotto eiaculatorio che si conclude in un gonopodio presente nello sternite dell'ottavo segmento addominale. La fecondazione č interna, avviene con un meccanismo copulatorio, il maschio deposita gli spermatozoi nella cavitą genitale della femmina per mezzo del gonopodio. Le uova prodotte sono rivestite da una membrana spessa, detta corion formatasi nell'ovario. Mentre le uova passano nell'ovidotto avviene la fecondazione. In molte specie (es. odonati) č presente un processo cuticolare attorno al gonoporo femminile formando un ovopositore che posiziona accuratamente le uova nel sito di incubazione, questo puņ essere lo stelo di una pianta o anche l'interno di un insetto ospite.
Come per gli altri artropodi, le uova degli unirami sono centrolecitiche con un abbondante materiale vitellino, il tipo di segmentazione, perņ, č meroblastica. Le uova degli unirami nelle fasi iniziali vanno incontro a segmentazione attraverso divisioni nucleari intralecitiche e successivamente si ha la migrazione di nuclei figli verso il periplasma (citoplasma periferico). Nel periplasma i nuclei continuano a dividersi fino alla saturazione e quindi iniziano a formarsi le membrane cellulari delimitando ogni nucleo. Dopo questi processi l'embrione č nella fase di periblastula costituita da una zona vitellina rivestita da uno strato di cellule o blastoderma. Da un lato della periblastula si forma un disco germinativo di cellule colonnari, tale disco č chiaramente distinguibile dal resto del blastoderma. A partire da specifiche zone del disco germinativo proliferano dei centri di formazione delle cellule endodermiche e mesodermiche: nello stadio di gastrula tali cellule si spostano verso l'interno e si dispongono al di sotto delle restanti cellule esterne che costituiranno l'ectoderma. Il mesoderma prolifera verso l'interno e forma il solco gastrico longitudinale, queste cellule che andranno a formare l'intestino rivestono la massa vitellina centrale e lentamente cominciano ad assorbirla; contemporaneamente si formano gli spazi celomatici. Progressivamente iniziano a delinearsi i segmenti corporei, ognuno dei quali presenta due tasche mesodermiche da cui emergeranno gli abbozzi delle appendici. Si formano i vari organi e tessuti, gli spazi celomatici si associano al blastocele andando cosģ a formare lo spazio emocelico. L'ectoderma si invagina parzialmente ai due apici dando origine alla bocca ed all'ano, da qui si formano l'intestino anteriore e posteriore (ectodermico stomodeale e proctodeale) che successivamente si congiungono all'intestino medio eutodermico. Questo č lo schema generale di sviluppo, i gruppi di unirami presentano molte variazioni soprattutto riguardo la determinazione delle cellule embrionali. Nei pauropodi e nei sinfili, ma anche in alcuni insetti come i collemboli si ha una segmentazione pił propriamente oloblastica in conseguenza di una riduzione secondaria della massa vitellina: i chilopodi ed alcuni insetti hanno un tipo di segmentazione intermedia tra totale e superficiale, il tuorlo si divide in regioni dette piramidi vitelline.
Nello sviluppo post-embrionale la maggior parte degli insetti cambia forma, si ha il processo di metamorfosi. Gli insetti procedono ad un certo numero di mute per accrescersi e raggiungere lo stadio adulto. 
Lo sviluppo puņ avvenire in tre modi diversi. METAMORFOSI COMPLETA. In questo tipo di sviluppo i processi fisiologici, di accrescimento, differenziamento e riproduzione sono separati e corrispondono rispettivamente allo stadio di larva, pupa e adulto. Le larve sono vermiformi con pezzi boccali masticatori, spesso vivono in ambienti completamente diversi dall'adulto. Quando cominciano a svilupparsi le ali all'interno del corpo della larva, questa inizia a formare un bozzolo intorno a sé e diviene una pupa o crisalide che rappresenta un condizione di quiescenza necessaria per superare l'inverno. Dopo la muta finale dal bozzolo emerge un adulto completamente sviluppato, le ali si distendono e si induriscono grazie alla circolazione dei fluidi corporei. L'adulto non subisce mute. Questo processo č proprio degli insetti appartenenti al gruppo Holometabola. METAMORFOSI GRADUALE O INCOMPLETA. Gli insetti con questo tipo di sviluppo, detto emimetabolo, presentano stadi giovanili che assomigliano ad adulti poiché hanno occhi composti, antenne ed appendici per la locomozione e l'alimentazione anche se non hanno ali funzionali e strutture sessuali. Gli stadi giovanili sono chiamati ninfe e presentano delle ali rudimentali. Le ninfe di odonati, effimere e libellule vivono in acqua, hanno le branchie e sono chiamate naiadi. SVILUPPO DIRETTO. Tale processo č tipico degli insetti atteri. Gli stadi giovanili sono molto simili agli adulti tranne che per le dimensioni e per le strutture sessuali.
Le strategie di sviluppo indiretto, olometabolo ed emimetabolo, sono selettivamente vantaggiosi, comportano infatti la separazione ecologica e cosģ facendo riducono la competizione intraspecifica.
Come gią accennato il processo di accrescimento, ed in particolare la metamorfosi, č controllato da ormoni. Gli organi endocrini funzionali nella crescita sono il cervello, le ghiandole protoraciche, i corpora cardiaca ed allata.
Il sistema nervoso contiene molte cellule neurosecretrici che producono l'ormone ecdisiotropina. Questo ormone viene trasportato dai liquidi emocelici fino alla ghiandola protoracica che cosģ riceve il segnale per il rilascio dell'ormone della muta o ecdisone. L'ecdisone permette l'eliminazione della vecchia cuticola o ecdisi. La muta č controllata anche da un'altra sostanza detta ormone giovanile che viene secreto dai corpora allata. Durante le prime mute la concentrazione dei due ormoni č elevata ogni muta da origine ad una larva pił grande. Verso gli ultimi stadi larvali la concentrazione di ormone giovanile č bassa e la larva muta e passa allo stadio di pupa o di ninfa. Durante l'ultima muta cessa la secrezione degli ormoni e si passa allo stadio adulto. Molti gruppi di insetti, soprattutto quelli che vivono in ambienti estremi, si sottopongono a partenogenesi stagionale.
|