|
The genus Metacyclops is widespread in tropical and temperate regions, being most prolific in European, African and South American regions, poorly known for North America and Australia. However, both the stygobiont and surface copepod fauna of Australia is still under study at present, and first results of our researches (Pesce et al., 1996a, 1996b; Pesce & De Laurentiis, 1996; De Laurentiis et al., 1997, in press) are revealing a rich assemblage of species in this area.
At present the genus includes 59 valid species and subspecies, 17 recorded in different groundwater habitats, such as wells, caves, anchialine habitats.
Metacyclops, as many other genera in the subfamily Cyclopinae [Muscocyclops Kiefer, 1937; Speocyclops Kiefer, 1937; Apocyclops Lindberg, 1942; Menzeliella Lindberg, 1954; Cochlacocyclops Kiefer, 1955 (= Metacyclops?); Goniocyclops Kiefer, 1955; Teratocyclops Plesa, 1981; Hesperocyclops Herbst, 1984; Fimbricyclops Reid, 1993; Graeteriella Brehm, 1926] has a proximal segment of leg 5, usually indicated by a remnant seta, partially or entirely fused to the somite, and a distinct distal segment with two appendages (Kiefer, 1927, 1929; Reid, in press). Among the above genera, Metacyclops is characterized by antennula 9-13, exceptionally 17-segmented, rami of the swimming legs 1 to 4 always 2-segmented, and free segment of leg 5 armed with one short spine and one, generally long, seta distally implanted.
Lindberg (1961) established two species-groups within the genus, viz. the minutus-group and the gracilis-group, embracing species characterized by one or two distal appendages on the endopod 3 of leg 4, respectively. In the same paper the Author divided the males of Metacyclops into two groups. viz. those with a leg 6 with three appendages, and those with the same leg bearing only two appendages. Herbst (1988, 1990) provided useful, updated World keys of the species.
The most recent review of the genus, with World distribution of species, was provided by Reid (1991). Later on, Rocha (1994) pointed attention to the presence of different patterns of integumental pores on the urosomites, as well as to other additional useful characters in the taxonomy of the genus. Pesce et al. (1996), as well, pointed out particular spinules pattern on the basipodite of the antennule, both in males and females,
According to the spine formula of the exopod 3 of legs 1 to 4, the following morphological groups can be pointed out, viz.:
| |
A - Species with spine formula: 3 4 4 3
B - Species with spine formula: 3 4 4 2
C - Species with spine formula: 3 4 3 3
D - Species with spine formula: 3 3 3 3
| | |
At present, the species M.grandis Kiefer, 1935 (Uruguay) cannot be placed in any of these groups since in the original description the author did not report the above spine formula.
Most species and subspecies belong to the first group (A). Within this group the species can be distinguished by numerous other characters, such as the number of articles of the female antennula, presence/absence of vestigial exopodal seta on the proximal article of the antenna, the armature of the coxa of legs 1 to 4, the number of distal appendages on the endopod 3 of leg 4, the length of furcal rami and the ratio between inner and outer apical furcal setae, the length ratio spine/seta on the exopod of female leg 5.
As regards this last character the following species can be separated since they are characterized by a very short seta and a rather long spine on the exopod of female leg 5, as well as they show a 11-segmented antennula and 2 apical spines on the distal segment of leg 4 endopod.
M.chelazzii Dumont, 1981 (Somalia, Africa)
M.micropus Kiefer, 1932 (Ivory Coast, Africa)
M.mutatus Herbst, 1988 (West Indies, Aruba)
The other species of this group are the following:
M.aequatorialis Dussart, 1977 (Ruanda, Africa)
M.agnitus Herbst, 1988 (West Indies, Barbados)
M.amoenus Mann, 1940 (Turkey)
M.botosaneanui Pesce, 1985 (West Indies, Bonaire)
M.brauni Herbst, 1962 (Brazil)
M.campestris Reid, 1987 (Brazil)
M.communis Lindberg, 1938 (India)
M.curtispinosus Dussart, 1984 (Venezuela)
M.dentatus (Plesa, 1981) (Cuba)
M.denticulatus Dussart & Frutos, 1985 (Argentine)
M.dianae Pesce, 1985 (West Indies, Haiti)
M.gasparoi Stoch, 1987 (Norheastern Italy)
M.geltrudae Galassi & Pesce, 1994 (Venezuela)
M.gracilis (Lilljeborg, 1853) (Europe, Africa, America, Asia)
M.grandispinifer Lindberg, 1940 (Turkey, Iran)
M.hannensis Defaye, 1992 (Senegal, Africa)
M.hartmanni Herbst, 1960 (Nicaragua) [syn. of M. laticornis ?]
M.hirsutus Rocha, 1994 (Brazil)
M.janstocki Herbst, 1990 (West Indies, Antigua)
M.laticornis (Lowndes, 1934) (Paraguay, Argentina, Brazil)
M.leptopus leptopus (Kiefer, 1927) (South America)
M.leptopus mucubajiensis Kiefer, 1956 (Soth America)
M.leptopus venezolanus Kiefer, 1956 (Venezuela)
M.leptopus totaensis Reid et al., 1990 (Colombia)
M.lusitanus Lindberg, 1961 (Portugal)
M.malayicus Kiefer, 1930 (Sumatra)
M.mendocinus (Wierzejski, 1892) (central and suthern America; Azores; Belgium ?)
M.mendocinus insulensis Defaye & Dussart, 1991 (Azores)
M.mendocinus venezolanus (Kiefer, 1956) (Venezuela, Brazil)
M.minutus (Claus, 1863) (Europe, Africa, Israel, Asia)
M.minutus senegalensis (Gauthier, 1951) (Senegal, Africa)
M.necessarius (Kiefer, 1926) (Africa)
M.oraemaris Rocha, 1994 (Brazil)
M.paludicola (Herbst, 1959) (Brazil)
M.pectiniatus Shen & Tai, 1964 (China)
M.planus (Gurney, 1909) (Europe, Africa, Asia)
M.postojnae Brancelj, 1987 (Yugoslavia)
M.problematicus Dumont, 1972 (Belgique)
M. prolatus (Kiefer, 1935) (Madagascar, Kenya)
M.pseudo-anceps Green, 1962 (Africa)
M.rudis Plesa, 1981 (Cuba)
M. cf. rudis Plesa, 1981 (Brazil, Cuba)
M.ryukyuensis Ishida, 1995 (Ryukyu Islans, Japan)
M.somalicus Dumont, 1981 (Somalia, Africa)
M.stammeri Kiefer, 1938 (South Italy, Montenegro) *
M.subdolus Kiefer, 1938 (Europe, Balearic Islands, Israel) **
M.subequalis Dussart, 1984 (Venezuela)
M.tredecimus (Lowndes, 1934) (South America)
M.trisetosus Herbst, 1957 (Yugoslavia)
M.tropicus Kiefer, 1932 (Africa) |
|
* The spine formula of this species is not reported in the original description by Kiefer; spine formula "3443" observed in specimens recently collected by Pesce et. al. from the type-locality (unpublished), and by Karanovic (1999) in material from Montenegro. The presence of this species in Corsica, Sicily and and Sardinia (Dussart & Defaye, 1985) is questionable (Karanovic, 1999). M. stammeri is one of the Tertiary relicts that survived Quaternary cyclic periods of ice ages in subterranean habitats. Its present distribution (fig. on the right) is most probably the consequence of the last ice age which drive it into two refuge areas [Salentine Peninsual, south Italy (1) and Skadar karstic valley (2)]. (after Karanovic,1999) | | | | | | 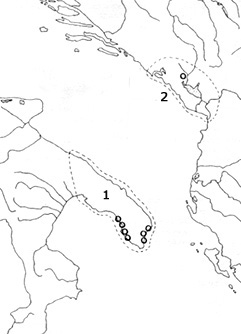 |
** The spine formula of this species is not reported in the original description by Kiefer; spine formula "3443" pointed out by Pesce (1978) in material from Greece and by Jaume (1990) in specimens from Balearic Islands; Por (1993) illustrated a leg 2 exopod 3 with only 3 spines in specimens of the same species from Israel ! Our recent examination of some specimens from the type locality confirmed a spine formula "3443" for this species (unpublished).
The second group (B) harbors only the recently described M. mortoni Pesce et al., 1996, limited to ground waters (wells, hyporheic) of Western Australia (Cape Range Peninsula).
The group "C" includes as well a single species, viz. M. cushae Reid, 1991 (Lousiana, U.S.A., Honduras, Brazil).
Five taxa fit the group "D", viz.: M. arnaudi (G.O. Sars, 1908), from Australia, M. arnaudi platypus Kiefer, 1967, also from Australia, M. margaretae (Lindberg, 1938), from India, M. monacanthus Kiefer, 1928, from New Zealand and M. trispinosus Dumond,1981, from Africa.
As regard M. margaretae, notwithstanding the different spine formula of the exopod 3 of legs 1 to 4, and other minor differences, Lindberg (1961) and successively Monchenko (1975), Dussart (1969) and Dussart & Defaye (1985) considered this species synonym of M. gracilis. However, in my opinion, and according to Defaye (in litt.), the two species can be considered as distinct taxa.

GALASSI D.M.P. & G.L. PESCE. 1994. Metacyclops geltrudae n.sp., a new cyclopid from ground waters of Venezuela (Copepoda, Cyclopidae). Crustaceana, 67 (3): 284-287.
ISHIDA T. 1995. A new species of Metacyclops (Crustacea, Copepoda, Cyclopoida) from Ishigaki Island, the Ryukyu Islands. Proc. Jap. Soc. Syst. Zool., 54:33-37.
KARANOVIC T. 1999. First record of Metacyclops stammeri Kiefer, 1938 from Balkan Peninsula. Spixiana, 22 (in press).
PESCE G. L., P. DE LAURENTIIS & W.F. HUMPHREYS. 1996. Copepods from ground waters of Western Australia. I. The genera Metacyclops, Mesocyclops, Microcyclops and Apocyclops (Crustacea Copepoda:Cyclopidae). Rec. West. Austr. Mus., 18: 67-76.
REID J. W. 1991. The genus Metacyclops (Copepoda:Cyclopoida) present in North America: M. cushae, new species, from Louisiana. Journ. Crustacean Biol., 11 (4): 639-646.
REID J.W., J.A. MOLINA AREVALO & M.M. FUKUSHIMA. 1990. Metacyclops leptopus totaensis new subspecies (Crustacea:Copepoda) from lago de Tota, Colombia. Proc.Biol. Soc. Washington, 103:674-680.
ROCHA C.E. F. 1994. New species of Metacyclops (Copepoda, Cyclopidae) from Brazil, with remarks on M. campestris. Zool. Script., 23: 133-146.

DEFAYE D. 1992. Metacyclops hannensis n. sp. (Crustacea, Copepoda,Cyclopoida), un cyclopide nouveau du Senegal. Rev. hydrobiol. trop., 25 (2):145-151.
HERBST H. 1988. Zwei neue Metacyclops (Crustacea Copepoda) von den Westindischen Inseln Barbados und Aruba: M. agnitus n.sp. und M. mutatus n.sp. sowie ein Bestimmungsschlussel fur das Genus. Bijd. Dierk., 58: 137-154.
HERBST H. 1990. Metacyclops janstocki n. sp. (Crustacea, Copepoda) von Antigua (Westindische, Inseln).
Beaufortia, 41 (11): 75-81.
LINDBERG K. 1961. Remarques sur le genre Metacyclops (Kiefer, 1927) et description d'un Metacyclops nouveau du Portugal. K, fysiogr. Sallsk. Lund. Forh., 31: 133-145.
REID J.W. 1987. The cyclopoid copepods of a wet campo marsh in central Brazil. Hydrobiologia, 153: 121-138.
REID J.W. 1990. Metacyclops leptopus totaensis, new subspecies (Crustacea:Copepoda) from Lago de Tota, Colombia. Proc. Biol. Soc. Wash., 103 (3): 674-680.
DEFAYE DANIELLE
Museum National d'Histoire Naturelle - Laboratoire de Zoologie (Arthropodes)
61, rue de Buffon, F-75005 Paris - France
e-mail: ddefaye@mnhn.fr
DE LAURENTIIS PAOLA
Dipartimento di Scienze Ambientali - Universitŕ di L'Aquila
Via Vetoio - 67100 - L'Aquila (Italy)
HERBST H.V.
Distelweg 1
4150 Krefeld 29 - Germany
ISHIDA TERUO
372 Irifunecho, Yoichimachi
Hokkaido, 046 Japan
KARANOVIC T.
Via Brescia, 3
84092 Bellizzi (SA) - Italy
PESCE GIUSEPPE L.
Dipartimento di Scienze Ambientali - Universitŕ di L'Aquila
Via Vetoio - 67100 - L'Aquila (Italy)
tel. 0039-862-433232; Fax: 0039-862-433205
e-mail: giuseppe.pesce@aquila.infn.it
http://www.univaq.it/~sc_amb/pesce.html
http://www.glpesce.net
REID JANET W.
Department of Invertebrate Zoology
MRC-163 National Museum of Natural History - Smithsonian Institution
Washington DC 20560- USA
tel. (202) 357-4674 fax (202) 357-3043
e-mail: 74024.1511@compuserve.com
ROCHA CARLOS E.
Departamento de Zoologia IBUSP
Caixa Postal 11461 - 05422-970 Sao Paulo, Brazil
Fax: 0055-11-8187416; e-mail: cefrocha@usp.br
 LINK TO THE COPEPOD WORLD AT SMITHSONIAN INSTITUTION LINK TO THE COPEPOD WORLD AT SMITHSONIAN INSTITUTION
Sincere thanks are due to D.Defaye and J.W.Reid
for useful suggestions and/or improvements Please, for comments and/or suggestions e-mail to: pesce@univaq.it |
[HOME] [BACK] |