 |
 CHECKLIST DELLE SPECIE DELLA FAUNA ITALIANA. COPEPODA Calderini ed. Italy - 1995 |

The copepods are the largest and most diversified group of crustaceans. At present they include over 14.000 species, 2.300 genera and 210 families, a surely underestimated number, inhabiting sea and continental waters, semiterrestrial habitats, or living in symbiotic relationships with other organisms. They are considered the most plentful multicellular group on the earth, outnumbering even the insects, which include more species, but fewer individuals! Particularly, the copepods are the dominant forms of the marine plankton and constitute the secondary producers in the marine environments and a fundamental step in the trophodinamics of the oceans.
During their long evolutionary history, starting in the Lower Cretaceous, copepods spread over all the continents, as well as they successfully colonized about all the available water habitats of the Planet, becoming well adapted or specialized to very different salinity regimes, from marine and hypersaline waters to continental freshwater bodies, and to a wide range of temperature from the polar to the hot springs waters. Copepods regularly live also in marine and freshwater sediments, in different groundwater substrates, in continental and anchialine caves (limestone caves), cenotes, ephemeral water bodies, marshes, wet campos, deep-sea hydrothermal vents, cold seeps and on plants as well. Moreover free-living cyclopoid and harpacticoid copepods may form an important component of cryptozoic fauna in moist forest litter (Reid, 1986; Fiers & Ghenne, 2000). Coastal, estuarine brackish waters harbor many copepod genera and species which can persist in these unfavourable habitats mainly as resting, diapausing eggs. Copepods can be found also in strange cryptic habitats, such as the small pools between the leaves of some tropical plants, crab burrows, tree holes and even discarded car tyres (!). Copepods, as many other aquatic organisms, can be easily transported, either actively or passively, often as resistant stages (Sars, 1901; Van de Velde 1984; Matsumura-Tundisi et al. 1990; Saunders et al., 1993; Reid & Reed 1994).
Neverthless, the astonishing success of this group of microcrustaceans is due to their symbiotic relationships with other organisms: in fact, they virtually can parasitize or be the intermediate hosts of all the animal groups, from sponges to vertebrates, including mammalians and man, and it is likely that the number of associated or parasitic taxa known today could represent only a small fraction of the living species, especially in marine waters. Copepods that parasitize fish skin and gills are serious pests of commercial importance in both marine and freshwater fish farms.
Some species of cyclopoid genera such as Acanthocyclops, Diacyclops, Macrocyclops and Mesocyclops are biological control agents for disease-bearing anopheline mosquitoes (Marten et al., 1994).
Stygobitic copepods originated from surface, marine and freshwater ancestors, and they reached different groundwater habitats mostly through interstitial and crevicular or karstic corridors. Typical groundwater harpacticoid and cyclopoid copepods (Graeteriella, Maraenobiotus, Moraria) seem to occupy large distribution areas, as a consequence of their greater dispersion capabilities as compared to those of macrofaunal subterranean elements (Fiers & Ghenne, 2000).
The systematics of copepods has been subjected to numerous revisions during the last decade and before: at present, according to Huys R.& G.A. Boxshall (1991), ten orders are recognized, half of which (Misophrioida, Monstrilloida, Mormonilloida, Siphonostomatoida, Poecilostomatoida) mostly contains exclusive symbiotic or parasitic species, the others (Platycopioida, Calanoida, Harpacticoida, Gelyelloida, Cyclopoida) as a rule include free-living forms. The following orders are represented in groundwater habitats : Platycopioida, Calanoida, Misophrioida, Gelyelloida, Harpacticoida and Cyclopoida.  [Morphology variation in the 10 Copepod orders; after Ho, 1990] The phylogenetic relationships of copepods have been debated in the last decades. Some authors proposed evolutionary schemes on account of the ecological radiation of copepods (Boxshall, 1986; Ho, 1994) , other authors pointed out cladistic approaches. Particularly Huys & Boxshall (1991) reconstructed the character states of the groundpattern of ten copepods orders. 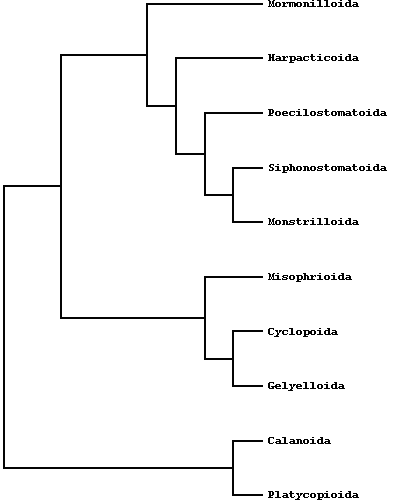 Calanoids, cyclopoids and harpacticoids show a remarkable ecological interest, since most species of these orders generally form the first link of the aquatic food chains, from the microscopic phytoplanktonic algae up to the fishes and mammalians. Recent researches, including those carried out by the Dipartimento di Scienze Ambientali of the University of L'Aquila (Italy), are pointing out an analogous importance for cyclopoid and harpacticoid species inhabiting both surface and underground fresh waters, and particularly the sediments between the superficial hyporheic zone and the rivers bottom, an interesting transitional system or ecotone between epigean and stygal waters. As a matter of fact, the contiguity with surface waters and the regular occurence of epigean elements in the hyporheic habitats let now the hydrobiologists to consider that a good estimation of rivers meiobenthic conditions must pass through a careful knowledge of the relative groundwater communities. In this last regard, an increasing number of harpacticoid and cyclopoid species are actually revealing their noteworthy importance as "pollution markers" in the environmental control of hyporheic systems and other aquatic habitats, such as lakes, springs, rivers and superficial ground waters. 
[Copepods Web Portal] [Feedback] [Previous Page]
|